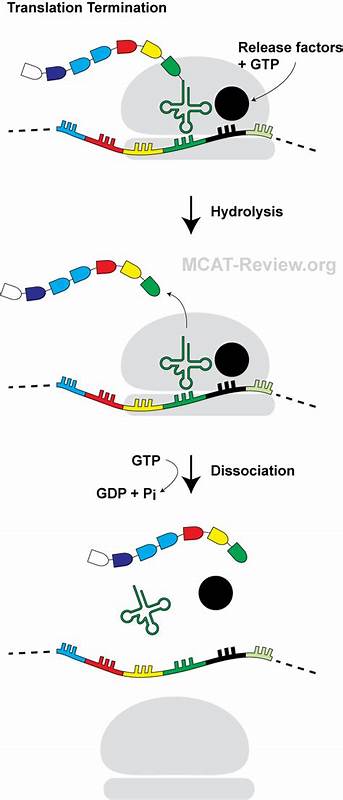
Die Proteinsynthese ist ein essenzieller Prozess in allen lebenden Zellen, bei dem die genetische Information von der mRNA in funktionelle Proteine übersetzt wird. In diesem komplexen Vorgang spielen die Ribosomen eine zentrale Rolle, indem sie Aminosäuren sequenziell zu Polypeptidketten verknüpfen. Die korrekte Beendigung der Translation ist ebenso wichtig wie ihr Start und Ablauf, denn erst durch die Translationstermination wird der neu synthetisierte Proteinkette freigesetzt und der Ribosomkomplex zur erneuten Verwendung vorbereitet. Ein bedeutsamer Schritt dabei ist die Erkennung von Stoppcodons durch die Freisetzungsfaktoren, welche die Peptidyl-tRNA-Handhabung katalysieren und die Abspaltung der Polypeptidkette ermöglichen. Bislang wurden verschiedene Mechanismen identifiziert, wie die ribosomale Translation reguliert oder sogar zum Stillstand gebracht werden kann, meist durch naszierende Peptide, die durch ihre Sequenz oder Faltung mit dem Ribosom interagieren.
Neueste Forschungen haben ein faszinierendes Mini-Haarpin-Peptid (PepNL) entdeckt, welches die Translationstermination auf eine ungewöhnliche und einzigartige Weise blockiert, und eröffnen damit neue Perspektiven für unser Verständnis der Proteinbiosynthese und deren Regulation. Das Mini-Haarpin-Peptid PepNL stammt aus der Bakterienart Escherichia coli und besitzt lediglich 14 Aminosäuren. Überraschenderweise führt die naszierende PepNL-Kette, wenn sie während der Translation im Ribosomstromkanal verharrt, zu einem spezifischen Abbruch der Terminationsphase an ihrem Stoppcodon, das in diesem Fall UGA ist. Im Gegensatz zu vielen anderen ribosomalen Arrestpeptiden benötigt PepNL jedoch keine zusätzlichen Was-kann-mich-Stoppen-Moleküle (arrest inducer), sondern wirkt allein durch seine Aminosäuresequenz. Die Struktur des PepNL-Peptids innerhalb des Ribosoms wurde mittels Kryo-Elektronenmikroskopie mit hoher Auflösung sichtbar gemacht und zeigt eine einzigartige Mini-Haarpin-Konformation.
Diese Faltung zeichnet sich dadurch aus, dass sich das N-Terminus-Fragment des Peptids entgegen der üblichen Verlaufsrichtung „nach hinten“ faltet, also in Richtung des Tunnel-Eingangs, anstatt zum Tunnel-Ausgang, was bisher so nicht beschrieben wurde. Das Besondere an der PepNL-Struktur ist, dass das Haarpin über mehrere hydrophobe Wechselwirkungen innerhalb des Peptids stabilisiert wird und gleichzeitig auch mit spezifischen Nukleotiden der 23S ribosomalen rRNA im Tunnel intermolekular interagiert. Diese Wechselwirkungen führen zu einer Verformung des nachfolgenden Peptidabschnitts, insbesondere am C-Terminus, wo es zu einem Sterik-Kollidieren mit dem für die Terminationsreaktion maßgeblichen katalytischen GGQ-Motiv des Freisetzungsfaktors RF2 kommt. In einem gesunden Terminierungsprozess schiebt sich das GGQ-Motiv von RF2 in das Peptidyl-Transferase-Zentrum (PTC) des Ribosoms und katalysiert die Hydrolyse der Esterbindung, die das Polypeptid an die tRNA bindet. Durch die Interaktion mit dem haarpinartigen PepNL wird diese Einbettung verhindert, was die Termination effektiv blockiert.
Diese Blockierung führt zu einer abrupten Beendigung der Proteinsynthese, da der Freisetzungsfaktor seine Funktion nicht ausführen kann und die naszierende Kette an der P-Stelle des Ribosoms festhängt. Experimental wurde dies sensationell durch eine Kombination aus Übersetzungs-Assays, Peptidyl-tRNA-Analyse mittels Massenspektrometrie und RNA-Fußabdruckmethoden gezeigt. Darüber hinaus konnte die Konstruktform des arrestierenden Peptids mit spezifischen Mutationen auf die für die Faltung wichtigsten Aminosäuren genauestens untersucht werden. Mutationen an den hydrophoben Residuen isolöslich mindestens vier (Ile3, Leu4, Ile8 und Tyr9) zerstörten das Haarpin, was das Arrestpotenzial von PepNL gänzlich aufhob. Diese präzise Korrespondenz zwischen Struktur und Funktion unterstreicht die hohe Spezifität und den eleganten Mechanismus dieses Peptids.
Ein weiterer bemerkenswerter Aspekt des PepNL vermittelt eine neue Sichtweise auf die Nature des Translation Terminationsmechanismus und dessen Regulation: Das PepNL-Peptid hemmt zwar RF2-vermittelte Termination, jedoch kann ein anderes molekulares Ereignis den Arrest aufheben. Bei Anwesenheit von ausreichend Tryptophan wird das Stopcodon von einem geladenen Tryptophanyl-tRNA (Trp-tRNATrp) gelesen, was zu einem sogenannten Stop-Codon-Read-Through führt. Das heißt, trotz des UGA-Stoppcodons wird eine Aminosäure eingefügt und die Translation wird über das ursprüngliche Stoppsignal hinaus fortgesetzt. Dieses Read-Through wirkt als natürlicher „Arrest-Inhibitor“ für die PepNL-abhängige Haltefunktion und zeigt, dass PepNL über eine neuartige Feedback-Regulation verfügt. Die Effizienz dieses Lesendurchgangs hängt offenbar auch vom zeitlichen Ablauf der Haarpin-Faltung ab – je schneller das Trp-tRNATrp an das Gering-Codon bindet, desto wahrscheinlicher wird der Arrest umgangen, bevor die Mini-Haarpinstruktur die Termination blockiert.
Dieses faszinierende Zusammenspiel eröffnet eine weitere Dimension für die Genexpressionskontrolle in Bakterien. Die Translation des pepNL sORFs reguliert die Expression des unmittelbar nachfolgenden pepN Gens in einem translation-coupled Mechanismus, welcher stark von der Peptidstruktur und der Zusammensetzung der zytosolischen Aminosäuren abhängt. PepN codiert eine Aminopeptidase, die eine zentrale Rolle bei der Protein- und Peptidverwertung sowie bei der Adaption an Umweltveränderungen spielt. Somit verbindet dieser Mechanismus die Translation mit dem metabolischen Status der Zelle. Die Entdeckung von PepNL als einzigartigem „mini-hairpin“ basierten Ribosom-Arrestpeptid stellt eine bedeutende Erweiterung des Wissens über ribosomale Regulationsmechanismen dar.
Während bisherige Studien vor allem auf Arrestpeptide fokussierten, die aufgrund externer Induktoren oder metabolischer Liganden ihre Wirkung entfalten, zeigt PepNL, dass allein die naszierende Peptidsequenz und ihre besondere Faltung suffice, um die Translationstermination zu steuern. Dadurch erhält die Bakterienzelle eine neue Art der fein abgestimmten Kontrolle über die Proteinproduktion in Abhängigkeit von der Verfügbarkeit bestimmter Aminosäuren. Die vielfältigen Wechselwirkungen, die das Mini-Haarpin im Ribosomtunnel eingeht, insbesondere die hydrophoben Bindungen zu den Basen der 23S rRNA, lassen darauf schließen, dass die ribosomale Tunnelumgebung eine dynamische Schnittstelle für translationale Regulationen ist. Frühere Studien mit anderen Arrestpeptiden wie TnaC, SpeFL oder MsrDL zeigten bereits, dass metabolisch induzierte Strukturänderungen im Ribosom die Translation verzögern können, aber die spezifische Rückfaltung des Naszierenden PepNL-Peptids entpuppt sich als eine einzigartige, strukturell stabile Konformation, die unabhängig von Liganden wirkt. Darüber hinaus wurden durch die hochauflösende Kryo-EM-Struktur signifikante Konformationsänderungen des Releasefaktors RF2 erkannt.
Insbesondere wurde eine Verschiebung des apikal gelegenen GGQ-Motivs beobachtet, die für die inaktive Konformation des Faktors verantwortlich ist. Die gemessene Verschiebung von etwa 16 Å verhindert den Zugang zum Peptidyl-Transferasezentrum und erklärt die Hemmung der Peptidfreisetzung. Diese Erkenntnisse liefern wertvolle mechanistische Einsichten, wie sogenannte Penultimate Residuen (residuale Aminosäuren direkt vor der C-Terminus-Sequenz) eine zentrale Rolle in der Feinregulierung der Termination spielen. Aus evolutionärer Perspektive wirft die Entdeckung von PepNL die spannende Frage auf, wie verbreitet solche miniaturisierten, selbstregulatorischen Arrestpeptide sind und welche weiteren zellulären Prozesse sie beeinflussen könnten. Das Screening von verschiedenen kleinen offenen Leseraster (sORFs) zeigte, dass neben PepNL weitere Kandidaten ähnliche Wirkungen haben können, was die Vielfalt und Komplexität der regulationellen Landschaft der Translation unterstreicht.
Somit wird angekündigt, dass in zukünftigen Studien nicht nur neue Peptide identifiziert, sondern auch deren spezifische Umgebungsfaktoren und Stimuli entschlüsselt werden müssen. Die methodische Kombination aus in vivo Overexpression, ribosomaler Fußabdruck-Analyse, hochauflösender Massenspektrometrie der peptidyl-tRNAs und der Kryo-Elektronenmikroskopie belegt eindrucksvoll die Funktionsweise von PepNL. Zusätzlich wurde anhand von gezielten genetischen Manipulationen der direkte Zusammenhang von Peptidstruktur und Translationsterminationsblockade validiert. Die Konkurrenz zwischen Freisetzungsfaktor und gelegentlicher Stop-Codon-Read-Through durch Trp-tRNATrp präsentiert sich als bisher nicht beschriebener Regulations-Loop. Zukünftige Forschung wird sich sicherlich der genauen zeitlichen Dynamik der Haarpin-Faltung und des Read-Through-Prozesses widmen sowie der Frage, wie zelluläre Signale diese Prozesse modifizieren.
Darüber hinaus könnte die gezielte Modellierung oder Entwicklung synthetischer Peptide, die ähnliche Mechanismen auslösen, neue therapeutische Interventionsmöglichkeiten eröffnen, insbesondere da bakterielle Translationsprozesse eine attraktive Zielstruktur für Antibiotikawirkstoffe darstellen. Zusammenfassend verdeutlicht die Integration von strukturellen, biochemischen und genetischen Befunden, dass das Mini-Haarpin-Peptid PepNL ein Paradigma für eine neue Art der translationellen Regulation darstellt. Seine Fähigkeit, die Translationstermination durch eine eigenständige Konformationsänderung des naszierenden Peptids und der Freisetzungsfaktoren zu blockieren, öffnet das Fenster zu einem noch nicht vollständig verstandenen Mechanismus der zellulären Genregulation. Die Erkenntnisse erweitern die Grundlagen unseres Verständnisses der Proteinbiosynthese und unterstreichen die Bedeutung kleiner Peptide bei der Regulation lebenswichtiger zellulärer Prozesse.








